基因编辑技术CRISPR/Cas9 “魔剪”一文通
时间:2022-03-11 14:13:12 热度:37.1℃ 作者:网络
基因编辑, “何方神圣”
基因编辑是一种对靶基因或转录产物进行敲除、插入和定点突变等精确修饰的基因工程技术,主要通过人工核酸酶实现对基因组的特定基因序列的敲除、插入或精确修饰。目前,主要有三大基因编辑技术,包括:锌指核酸酶 (Zinc finger nucleases; ZFNs) 技术,转录激活因子样效应物核酸酶 (transcription activator-like (TAL) effector nucleases; TALENs) 技术和CRISPR/Cas9技术。ZFNs是最较早用于基因组编辑的人工合成的限制性内切酶,该酶是异源二聚体,包含DNA结合锌指蛋白(ZFP)结构域和非特异性FokI核酸酶结构域。DNA切割域的FokI核酸酶必须二聚化以切割DNA。该技术已被用于修饰各种生物体内的内源性基因。与ZFNs的模块化结构一样,TALENs的羧基末端也含有FokI核酸酶结构域,它借助TALEs(来源于植物致病性黄单胞菌属细菌)来识别特异性DNA碱基对。相较于ZFNs技术,其优势在于可以定点识别靶基因,从而使得基因编辑更加准确高效,其脱靶效应以及细胞毒性也得到了显著改善。
图1. 不同基因编辑手段的对比
2020年,两位女性科学家Emmanuelle Charpentier和Jennifer A. Doudna因发现CRISPR/Cas9基因剪刀,而获得2020年诺贝尔化学奖。
CRISPR/Cas系统由一小段RNA和一种高效的DNA切割酶(Cas核酸酶)组成的系统,该技术不像TALENs技术和ZFNs技术是依赖于蛋白与靶基因之间的识别,而是由sgRNA和靶基因之间形成复合物,从而完成特定基因序列的编辑。
CRISPR/Cas9技术切割效率相对较高且操作简单,并且可以同时进行多位点的编辑。总之,三种不同的基因编辑技术都有其各自的特点。今天小M的“重头戏”,CRISPR/Cas9。
CRISPR/Cas9的调控机制
在细菌及古细菌中,CRISPR系统共分成3类,其中I类和Ⅲ类需要多种CRISPR相关蛋白(Cas蛋白)共同发挥作用。而来自Streptococcus pyogenes的Ⅱ型系统只需要一种Cas蛋白(Cas9)即可发挥核酸内切酶活性。因此,CRISPR/Cas9系统应用最为广泛。CRISPR/Cas9系统已经成功应用于植物、细菌、酵母、鱼类及哺乳动物细胞。
■第一步:捕获外源DNA
Cas9蛋白包含两个核酸酶结构域:切割非互补DNA链的RuvC结构域和切割互补DNA链的HNH结构域(如图2a)。
噬菌体等外源DNA入侵时,CAS蛋白复合物通常识别3'端含NGG的前间隔序列邻近基序(PAM)区域。然后,侵入的噬菌体或质粒释放的短DNA片段(称为Protospacer),插入宿主CRISPR位点中(由重复序列隔开)。
■第二步:crRNA合成
CRISPR序列转录,形成前体CRISPRRNA(pre-crRNA)。Cas9及RNaseIII在tracrRNA与pre-crRNA上的重复序列配对形成双链RNA的条件下,对pre-crRNA进行剪切,形成成熟的tracrRNA-crRNA双链RNA(即sgRNA)。Cas9核酸酶和sgRNA形成Cas9核糖核蛋白(RNP)。
■第三步:靶向干扰
当外源DNA再次进入细胞时,Cas9蛋白携带sgRNA,去识别外源DNA的Protospacer(前间隔序列),并与之结合,通过Cas9解旋酶和核酸酶对靶基因进行剪切。造成靶基因DNA的双链断裂(DSB),从而达到干扰靶基因表达的目的,在修复断裂同时引入基因敲除或敲入。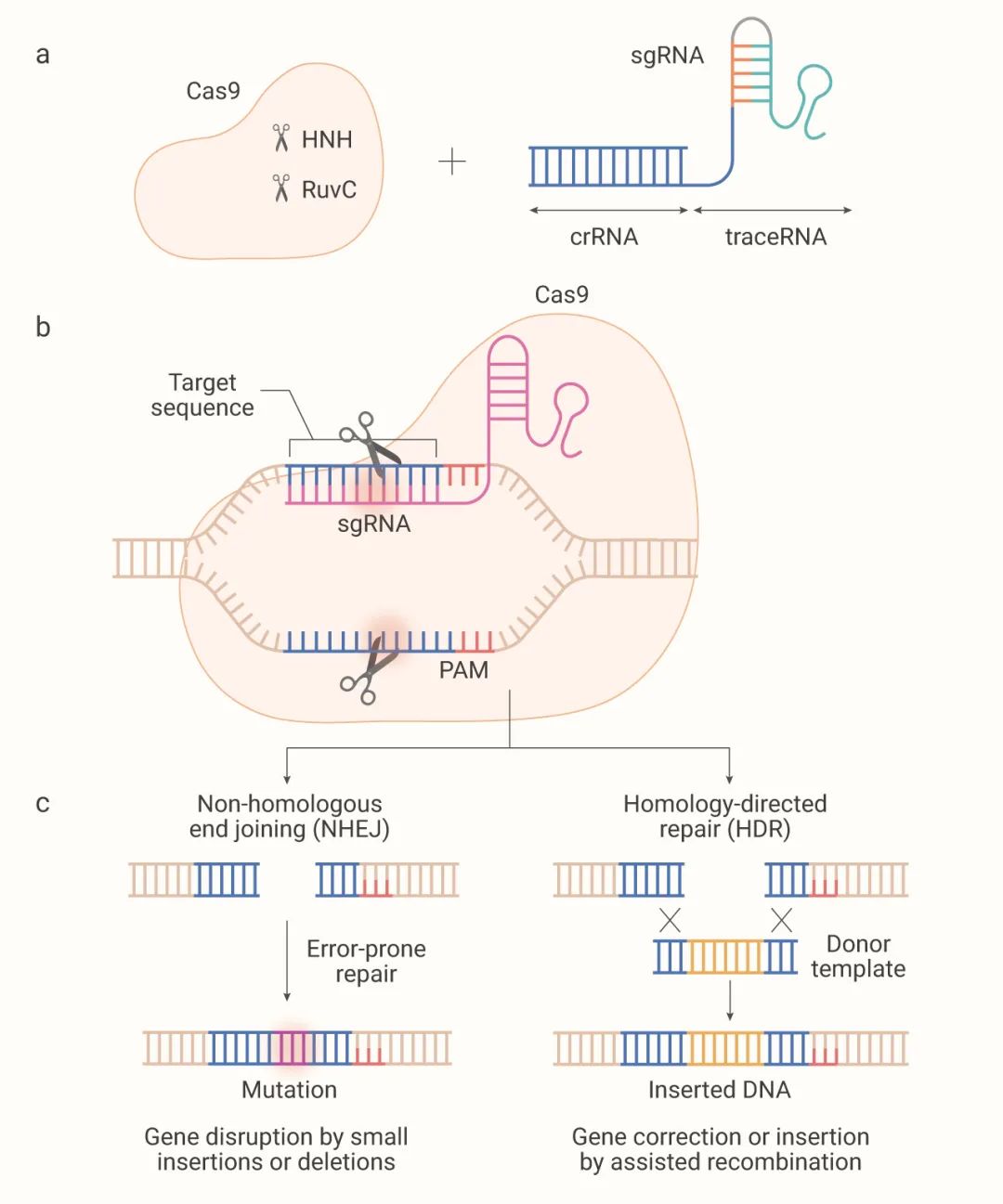
图2. 通过非同源末端连接 (NHEJ) 或同源定向修复 (HDR) 内源性修复双链DNA断裂[2]
综上,CRISPR技术主要是利用位点特异Cas核酸酶在基因组靶位点处引入DNADSB,再经细胞自身的非同源末端连接(NHEJ)或同源重组修复(HDR)对DSB进行修复,最终实现目标基因敲除和碱基编辑等基因组遗传修饰。
CRISPR/Cas9的小分子调控策略
CRISPR技术在疾病治疗、基因功能调控、药物研发等多个方面具有广阔的应用前景,但也存在脱靶、基因毒性等副作用问题。由于Cas9蛋白和sgRNA在其自身活性、识别位点及结合能力等方面的不同特性,因此在应用中可以通过对Cas9蛋白酶以及与靶DNA的结合进行有效的调控。目前,如:遗传调节、小分子激活剂、小分子抑制剂、生物响应性输送载体以及CRISPR/Cas9系统的光/热/超声/磁激活等方法已被研究开发。
■小分子激活剂控制Cas9活性的策略
可通过添加小分子来控制Cas9的构象变化,来实现对Cas9蛋白活性的时空控制。
案例1:当内含肽插入Cas9蛋白的不同位点,Cas9核酸酶失活;添加4-HT(4-hydroxytamoxifen),通过构象变化和自切割反应去除内含肽,可重新激活Cas9蛋白(图3a)。根据内含肽的插入位点,Cas9的激活效率3倍到10倍不等,与野生型Cas9相比,这种调控策略可增加开/脱靶效应比率。
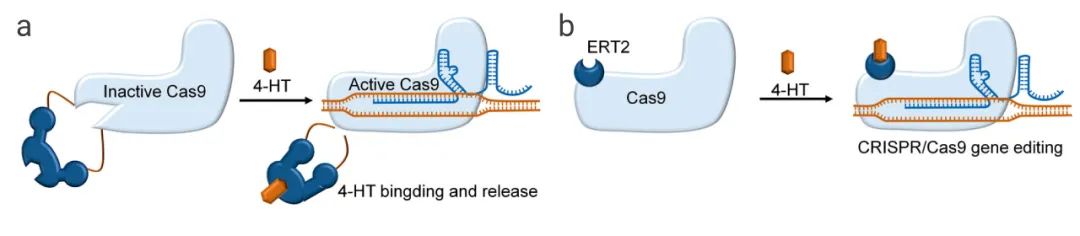
图3. 小分子激活剂控制Cas9活性的策略[9]
a:通过4-HT结合,恢复失活的Cas9活性;b:Cas9和雌激素受体(ERT2)的融合被分离在细胞质中,通过添加4-HT使得融合物进入细胞核,形成Cas9/sgRNA复合物
案例2:基于化学诱导的Cas9蛋白分裂片段的二聚化。Zetsche等人设计了不同的分裂位点(Arg535和Glu573)生成分裂Cas9(split-Cas9)蛋白,同时产生C端和N端Cas9片段(可分别与FK506结合蛋白(FKBP)和FKBP雷帕霉素结合结构域(FRB)结合)。这种方法通过雷帕霉素诱导的异源二聚作用实现了split-Cas9的条件重构和激活。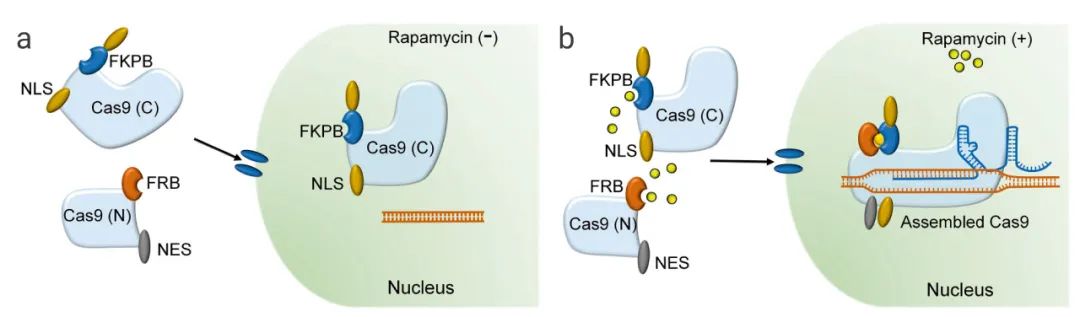
图4.小分子激活剂控制Cas9活性的策略[9]
■小分子抑制剂控制Cas9活性的策略
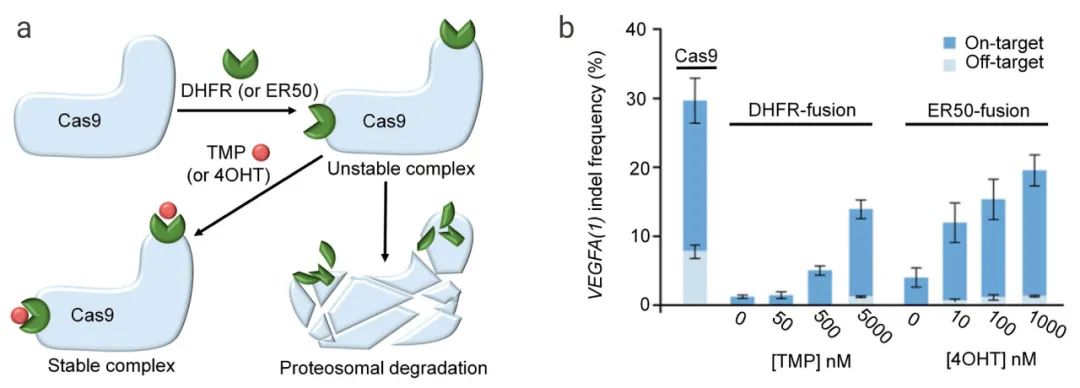
由于Cas9活性的升高和持续可能会导致脱靶效应、染色体易位和遗传毒性,因此在目标编辑后,Cas9核酸酶活性必须迅速限制在一个狭窄的时间范围内。
如下图5所示,DHFR(ER50)是一个不稳定的结构域,可快速靶向Cas9蛋白进行蛋白酶体介导的Cas9降解,但添加小分子抑制剂甲氧苄氨嘧啶(TMP)或4-OHT后可以使其稳定。用Cas9-DHFR或Cas9-ER50系统编辑VEGFA基因时,用不同剂量的TMP或4OHT剂量依赖性控制靶向VEGFA基因的复合物Cas9-DHFR(ERR50)的靶向和非靶向活性。